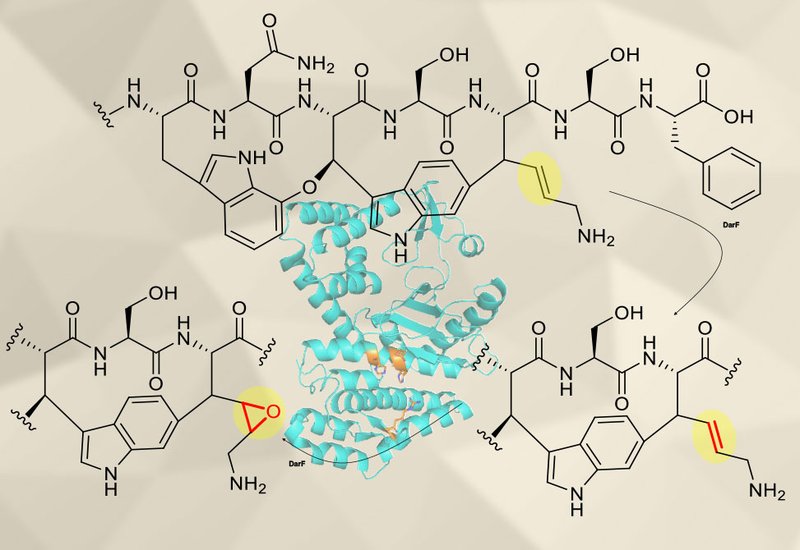
Darobactin A is a ribosomally synthesized and post-translationally modified peptide, RiPP, antibiotic that targets BamA, the essential outer membrane protein of Gram-negative bacteria. Its biosynthetic gene cluster encodes several tailoring enzymes, and the broader family of natural products assembled by the radical S-adenosylmethionine enzyme DarE has been named the daropeptides. When researchers isolated darobactin A and three structural relatives from the marine bacterium Pseudoalteromonas luteoviolacea H33 in 2023, the halogenase responsible for bromination was characterized, but the enzymatic origin of one derivative, dehydrodarobactin A, remained unexplained. The gene product DarF had been proposed as a self-resistance protease that degrades the antibiotic; its true biosynthetic role was unknown.
Researchers in the Phan Lab at the Latvian Institute of Organic Synthesis, published in Org. Biomol. Chem., re-examined DarF using a reconstituted in vitro system informed by the recent characterization of related HExxH enzymes. Earlier work on DarF had tested zinc as a cofactor, consistent with the metalloprotease hypothesis. The team instead supplied Fe2+, α-ketoglutarate, and ascorbate, cofactors appropriate for α-ketoglutarate-dependent non-heme iron dioxygenases. A modified full-length precursor peptide substrate was prepared by co-expressing the precursor DarA together with the crosslinking enzyme DarE in Escherichia coli, generating the bicyclophane-bearing scaffold required for downstream tailoring.
Incubation of the modified DarA substrate with purified His6-DarF produced two new species absent in heat-inactivated controls. Tandem mass spectrometry localized a +10 Da mass shift to the core peptide in the first product and a +26 Da shift in the second. Time-course and enzyme concentration-dependent assays established the reaction order: the substrate converts first to the +10 Da dehydrogenated species, which then accumulates the additional +16 Da to yield the +26 Da epoxidized product. Cofactor-dependence experiments confirmed that both Fe2+ and α-ketoglutarate are required for activity, placing DarF firmly within the α-ketoglutarate-dependent oxidase family despite its HExxH sequence motif. AlphaFold3-predicted structural comparison via the DALI server identified the cyclophane hydroxylase ChlH as the closest structural neighbor, and superimposition with the ChlH crystal structure revealed that a single arginine residue, Arg282, appears to substitute for the PWRxxxRP motif that coordinates α-ketoglutarate in the related enzymes.
To complement the in vitro data, the team analyzed culture extracts of P. luteoviolacea H33 by UHPLC-MS and confirmed the presence of darobactin A, dehydrodarobactin A, and epoxidized darobactin A in the native producer. The dehydrogenated product matches the previously NMR-verified dehydrodarobactin A, while the epoxidized species is predicted from mass data without NMR confirmation. Phylogenetic analysis shows DarF sits outside the clade containing the HExxH hydroxylases ChlH, MscH, and SjiH, consistent with its divergent chemistry. Among α-ketoglutarate-dependent enzymes acting in RiPP biosynthesis, only the dioxygenase ColD had previously been reported to perform sequential dehydrogenation–epoxidation; DarF is the second such example, and only three enzymes across ribosomal and non-ribosomal peptide pathways, ColD, AsqJ, and BcmB, had been described with this two-step oxidative logic before this study.
Reassigning DarF from protease to oxidative tailoring enzyme resolves a missing biosynthetic step in the darobactin pathway and expands the known chemical repertoire of HExxH-motif proteins. The authors note that the protease activity observed in earlier studies, which used leaderless darobactin A as substrate, is not ruled out; a closely related enzyme, PflC, has independently been shown to switch between oxidative and proteolytic functions depending on whether the leader peptide is present. For synthetic biology and natural product engineering, the identification of DarF as the enzyme responsible for dehydrodarobactin A provides a genetic handle for generating new daropeptide variants, including those bearing the cyclophane-ring dehydroamino acids found so far exclusively in this compound class.